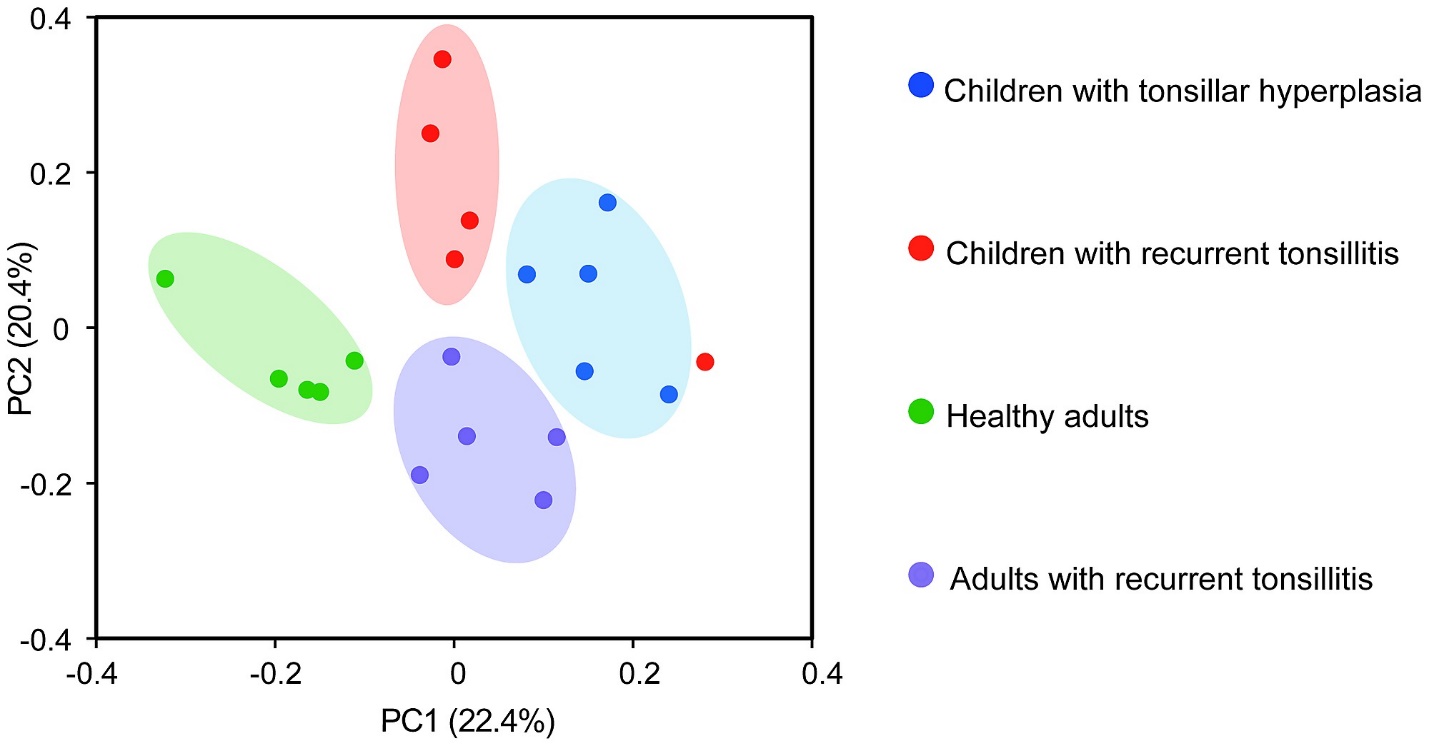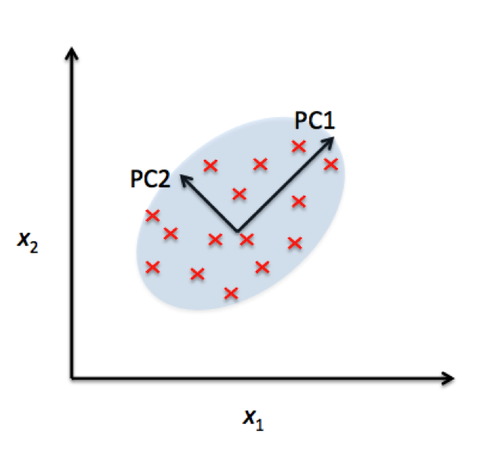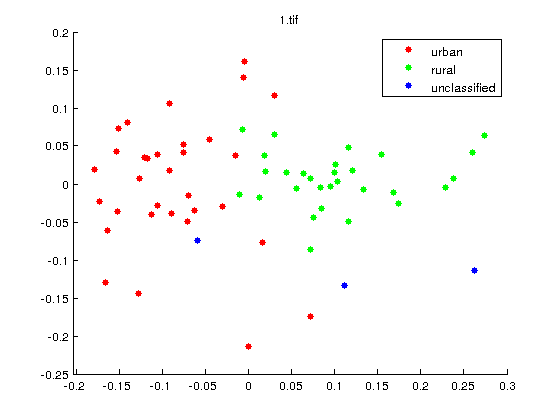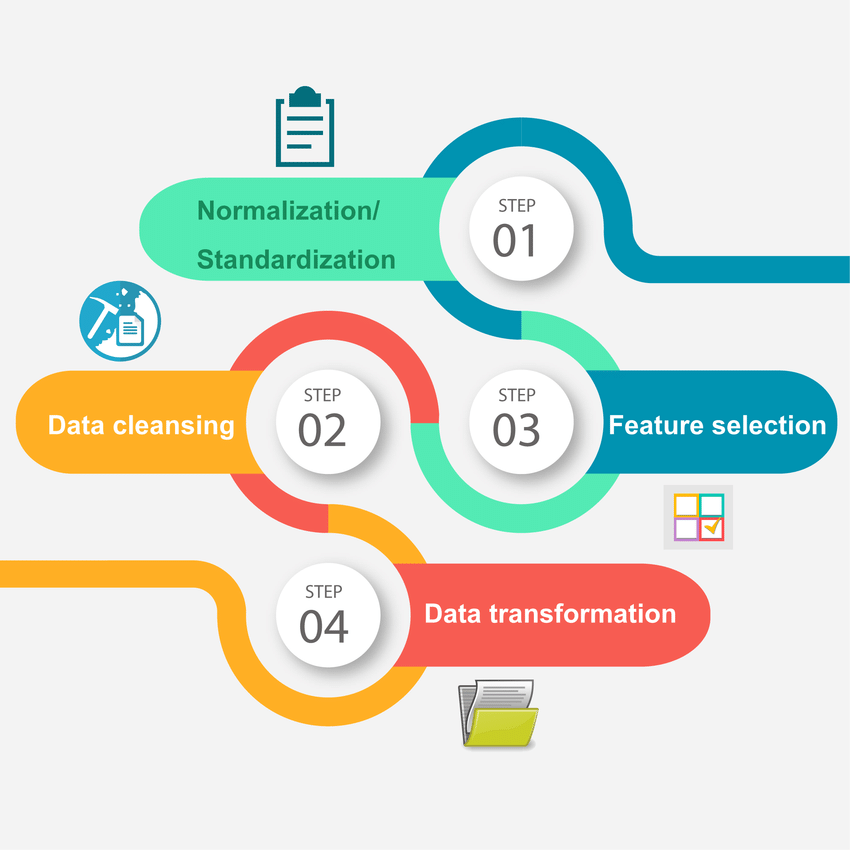
What are the differences between Labelled and Unlabelled Data?
Labeled data is a group of samples that have been marked with one or more labels. Labeling typically takes a set of unlabeled data and expands each piece of that unlabeled data with meaningful tags that are informative.
Unlabeled data is a description for pieces of data that have not been tagged with labels identifying characteristics, properties or classifications. Unlabeled data is typically used in various forms of machine learning.
Share: